
Inflammation is a natural immune response to neutralize and clear foreign bodies and damaged cells in a biological organism (Ilich, Kelly, Kim, & Spicer, 2014). The immune response can have several etiologies such as overconsumption of omega-6 rich foods, food sensitivities, poor sleep, stress, toxic environments, and intense physical activity (Ruiz-Nunez, Pruimboom, Dijck-Brouwer, & Muskiet, 2013).
However, viewing the immune response as “good” or “bad” might be an oversimplified and fallacious generalization. Instead of anthropomorphizing the inflammatory response, one might view it as a physiological event that exists along a spectrum; acute exposures to stimuli, up to chronic exposures to stimuli. The following sections will explore inflammation along the aforementioned spectrum, as a means of contextualizing the immune response, as well as understanding when it is beneficial, and when it is not.

Adaptation is a reaction to the environment, enabling humans to survive (Ruez-Nunez et al., 2013). The ideal amount of stressors (i.e., exercise, sun exposure) will yield an immune response that improves an individual’s resilience to the same stressor when re-exposed. However, adaptation exists along a bell curve, in that a dose-response relationship exists between a stressor and immune response; acute exposure can yield favorable adaptations, where chronic responses can develop pathophysiological conditions (Radak, Chung, Koltai, Taylor, & Goto, 2008). The optimal balance of conditions to develop an adaptation to a stressor, which improves health and performance, is known as hormesis (Radak et al., 2008).
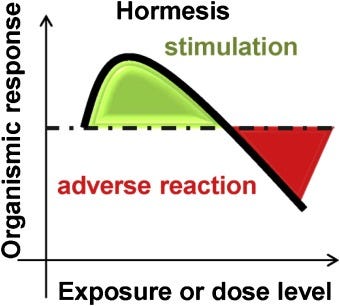
Hormesis is characterized by exposure to a stressor followed by adaptation. Specifically, two variables are central to hormesis; sufficient exposure to the stressor, and adequate recovery (Radak et al., 2008). Exercise, for example, when moderately intense and brief, allows an opportunity for adaptation, when followed by adequate rest periods. Several positive adaptations occur from mechanical and physiological loading (stressors) whereby there is disruption of the myofilament network, damage to the sarcomeres, splitting of the z-band, and increased intramuscular pressure (Radak et al., 2008).
The damage activates a brief inflammatory process, using signaling molecules (i.e., NF-κB) to produce inflammatory molecules (i.e., proteases), which help breakdown damaged proteins as part of the re-building process (Radak et al., 2008). The following sections will explore the inflammatory response more deeply, as well as its relationship to hormesis.
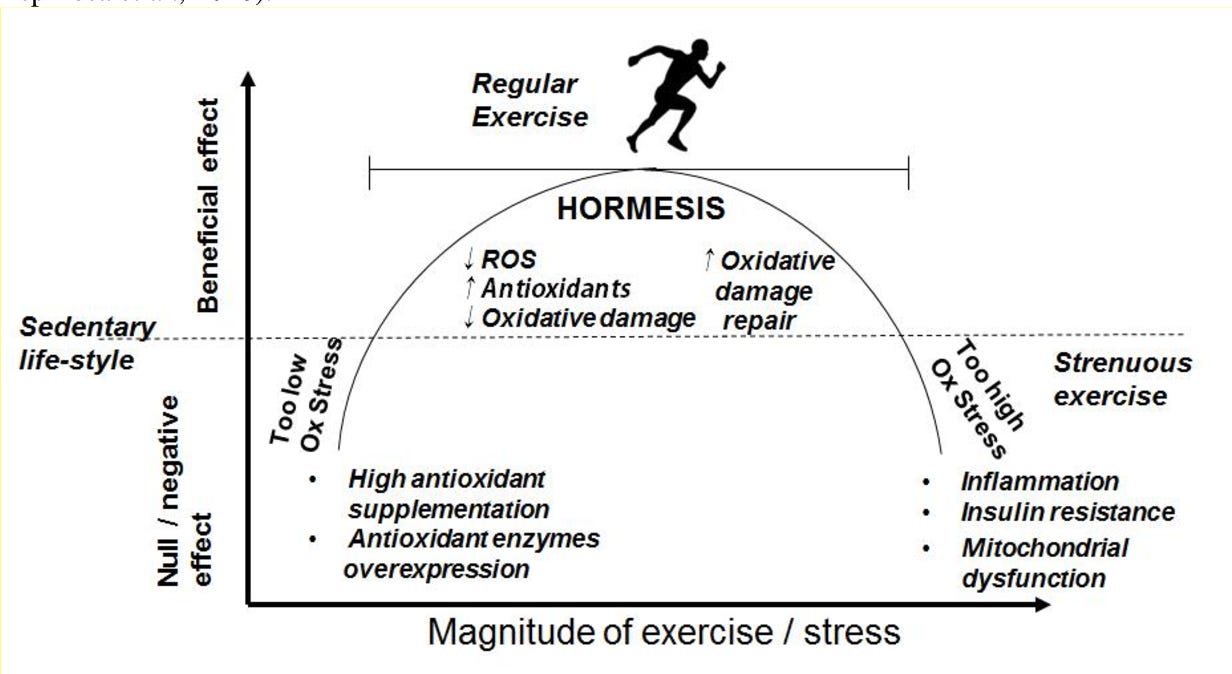
Adequate exercise prescription induces a brief proliferation of reactive oxygen species (ROS). ROS are a part of normal cellular function and plays a role in cell signaling, including apoptosis, gene expression, and the activation of cell signaling cascades (Radak et al., 2008). When ROS appear from brief and moderately intense exercise, it causes the immune system to respond by increasing antioxidant activity and damage repair enzymes. Thus, well-structured exercise programs improve the anti-oxidant defense system such as superoxide dismutase (SOD), catalase, and glutathione peroxidase (GSH); all of which suppress pro-inflammatory mediators and strengthen the immune system (Radak et al., 2008).
Such a response improves an individual’s resilience to the same exercise intensity and ROS when re-exposed. However, when the intensity and frequency of exercise is excessive, with little recovery time, the opportunity for meaningful hormesis is lost; overloaded mechanical stressors disintegrate sarcomeres and can tear tendo-muscular junctions, causing a large immune response, which induces apoptosis (i.e. programmed cell death) and necrosis (i.e., tissue death). Thus, awareness of training duration and intensity becomes paramount to achieve hormesis.
A balance of ideal inflammation also exists within the realm of nutrition, and is greatly influenced by fatty acid consumption and fatty acid type. Omega-6 and omega-3 polyunsaturated fatty acids (PUFAs) undergo a biochemical change once absorbed. The metabolic products of both PUFAs include eicosanoids; signaling molecules responsible for the control of immune and inflammatory reactions in the body. Pro-inflammatory Omega-6 eicosanoids include prostaglandin E2, leukotriene B4, and thromboxane A2.
Eicosanoids are generated by the enzymes lipoxygenase-5 and cyclooxygenase-2 (Ilich et al., 2014). Omega-3 eicosanoids, which are anti-inflammatory in nature, include prostaglandin E3, leukotriene B5, resolvins and protectins. Here, anti-inflammatory eicosanoids also use lipoxygenase-5 and cyclooxygenase-2 (Ilich et al., 2014).
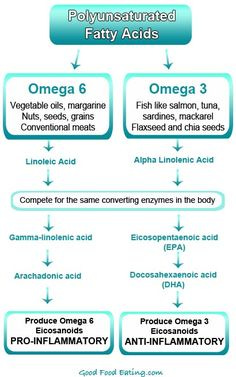
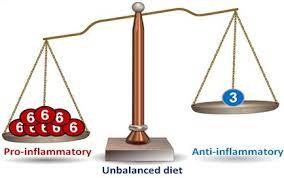
Of particular relevance and interest are the shared enzymes between the two PUFAs (i.e., lipoxygenase-5 and cyclooxygenase-2); both PUFAs compete for use of lipoxygenase-5 and cyclooxygenase-2. Thus, if more omega-6 fatty acids are present in the diet than omega-3 fatty acids, more pro-inflammatory signaling molecules are created, and larger levels of systemic inflammation will occur. If chronic consumption of high omega-6 / omega-3 ratios continue, chronic systemic inflammation ensues (Ilich et al., 2014).
An inflammatory response also produces cytokines such as tumor necrosis factor alpha, interleukin-6, and interferon-gamma; proteins important in cell signaling and immune responses (Ilich et al., 2014). Low-grade chronic inflammation (LGCI) from overconsumption of omega-6 PUFAs perpetuates the production and levels of these pro-inflammatory signaling molecules. Thus, although omega-6 PUFAs encourage inflammation, which is a necessary immune response to acute trauma and infections, chronic overconsumption of omega-6 PUFAs has deleterious effects that are associated with many Western diseases, commonly known as metabolic syndrome (Ruez-Nunez et al., 2013).
Inflammation has a time and a place; acute trauma and infections require the expeditious response of a strong immune system to help guarantee adaptation, and ultimately, survival. However, such a response must be brief and specific to circumvent collateral damage from the overproduction of ROS. In conclusion, if individuals embrace the principles of hormesis in an intelligent and pragmatic fashion, there exists an opportunity to achieve health, performance, and longevity in a way that optimizes immune system function, and adaptation.
References
Ilich, J.Z., Kelly, O.J., Kim, Y., & Spicer, M.T. (2014). Low-grade chronic inflammation perpetuated by modern diet as a promoter of obesity and osteoporosis. Archives of Industrial Hygiene and Toxicology, 65(2), 139-148.
Radak, Z., Chung, H.Y., Koltai, E., Taylor, A.W., & Goto, S. (2008). Exercise, oxidative stress, and hormesis. Aging Research Reviews, 7(1), 34-42.
Ruiz-Nunez, B., Pruimboom, L., Dijck-Brouwer, D.A., & Muskiet, F. (2013). Lifestyle and nutritional imbalances associated with Western diseases: Causes and consequences of chronic systemic low-grade inflammation in an evolutionary context. Journal of Nutritional Biochemistry, 24(7), 1183-1201.
-Michael McIsaac